
随着養殖業的快速發展,人工(gōng)配合飼料的使用量逐年增加,導緻飼料資(zī)源日趨緊張。因此,傳統飼料資(zī)源的高效利用及新型飼料資(zī)源的開(kāi)發就顯得尤爲重要。爲了解決上述問題,近年來,人們逐漸将目光轉移到發酵飼料這個新型飼料資(zī)源上。發酵飼料是以配合飼料爲發酵底物(wù),選擇接種特定的微生(shēng)物(wù),人工(gōng)控制溫度、水分(fēn)、需氧量等條件,通過微生(shēng)物(wù)自身繁殖和代謝來生(shēng)産的富含高活性益生(shēng)菌及其代謝産物(wù)的飼料。與傳統飼料相比,發酵飼料具有抗營養因子含量低、營養價值高、小(xiǎo)分(fēn)子蛋白(bái)和肽類含量高等優點。目前,發酵飼料已用于凡納濱對蝦( Litopenaeus vannamei) 、黑鲷( Acanthopagrus schlegelii) 、斑點叉尾 ( Ictalurus punctatus) 、草魚( Ctenopharyngodon idellu) 等的研究。但相對而言,其在水産中(zhōng)的應用研究尚較少,相關作用機制和效果還有待進一(yī)步探究。此外(wài),不同發酵飼料由于使用的微生(shēng)物(wù)菌株及發酵工(gōng)藝不同,其應用效果差異較大(dà),較難獲得穩定、一(yī)緻的效果。
微生(shēng)物(wù)發酵是發酵飼料加工(gōng)過程中(zhōng)的一(yī)個重要環節,在此過程中(zhōng)往往會産生(shēng)一(yī)定的酸類物(wù)質。因此,過量使用發酵飼料往往會導緻水産動物(wù)腸道 pH 值下(xià)降、肌肉中(zhōng)酸性物(wù)質的沉積并對其造成應激,進而其腸道損傷、肌肉品質下(xià)降和免疫機能受損,最終影響動物(wù)的生(shēng)長性能、抗病力和養殖效益。鑒于此,在評估發酵飼料在水産養殖中(zhōng)的應用潛力時,考察其對動物(wù)腸道消化吸收功能、肌肉品質及免疫機能的影響顯得尤爲重要。
因此,爲了對發酵飼料在水産養殖中(zhōng)的應用效果進行科學評估,本試驗以我(wǒ)(wǒ)國重要經濟魚類———鯉魚( Cyprinus carpio) 爲研究對象,從生(shēng)長性能、體(tǐ)組成、腸道消化酶活性、肌肉品質及免疫機能等方面入手,探讨了發酵飼料對鯉魚幼魚生(shēng)長性能及生(shēng)理功能的影響,以期爲發酵飼料在水産飼料中(zhōng)的應用和新型水産飼料資(zī)源的開(kāi)發提供技術依據。
1 材料與方法
1.1 試驗設計與飼料組成
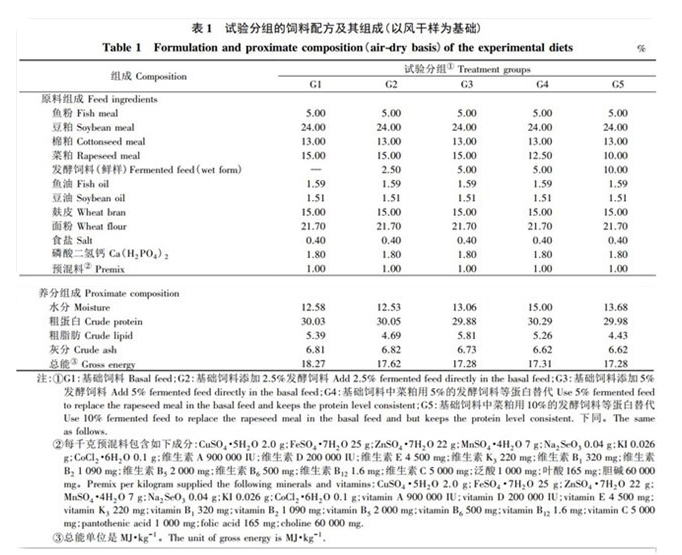
本試驗中(zhōng)所用的發酵飼料和益生(shēng)菌由南(nán)京寶輝生(shēng)物(wù)飼料有限公司提供,其他原料均由徐州正昌飼料有限公司提供。發酵飼料是以豆粕、菜粕、次粉、玉米等作爲底物(wù),由酵母菌、芽胞杆菌和乳酸菌作爲發酵菌種,并采用好氧和厭(yàn)氧方法進行發酵而成。其成分(fēn)大(dà)緻如下(xià)( 本文中(zhōng)“%”除特别注明外(wài),均表示質量分(fēn)數) : 水分(fēn) 42%、粗蛋白(bái) 18%、粗灰分(fēn) 3%、粗脂肪 1.3%、粗纖維 6%。本試驗共配制 5 組試驗飼料: 基礎飼料( 對照組, G1) 以魚粉、豆粕、菜粕和棉粕爲主要蛋白(bái)源,以麸皮和面粉爲糖源,魚油和豆油等比例添加爲脂肪源。G2 和 G3 組的飼料是在基礎飼料中(zhōng)添加 2.5%和 5% 的發酵飼料( 鮮樣) 。此外(wài),設定 2 個替代組: 以 5% 與 10% 的發酵飼料( 鮮樣) 等蛋白(bái)替代對照組的菜粕,即 G4 和 G5 組。試驗飼料的配方及營養成分(fēn)含量見表 1。将原料粉碎後過 60 目篩,然後與磷酸二氫鈣、預混料和食鹽等微量原料進行預混合,再将其加入其他粉碎原料中(zhōng)逐級充分(fēn)混勻,加入油脂和适量水後用小(xiǎo)型制粒機制成粒徑 2 mm 的顆粒飼料,室溫避光晾幹後保存于-20 ℃ 冰箱冷藏備用。
1.2 試驗魚與養殖管理
養殖試驗在南(nán)京農業大(dà)學浦口試驗基地的戶外(wài)網箱内進行,試驗所用鯉魚由中(zhōng)國水産科學研究院淡水漁業研究中(zhōng)心提供。正式試驗開(kāi)始前,将鯉魚暫養于網箱中(zhōng),期間投喂商(shāng)品飼料使其适應試驗環境。馴化 1 周後,将 300 尾體(tǐ)格健壯、規格整齊、初始體(tǐ)質量爲( 25.23±0.06) g 的鯉魚随機分(fēn)成 5 組,每組 4 個重複,每個重複 15 尾魚,置于一(yī)個網箱中(zhōng)。養殖試驗的 20 個網箱( 規格爲 1.0 m×1.0 m×1.5 m) 在同一(yī)池塘中(zhōng)進行,養殖期爲 10 周。每組飼喂 1 種飼料,每日定時飽食投喂 3 次( 07: 00、11: 30 和 16: 00) ,每日觀察并記錄魚攝食及死亡情況。試驗期間水溫 25 ~ 35 ℃ , pH 值爲 7.0 ~ 7.5,溶解氧含量大(dà)于 5.0 mg·L-1,氨氮含量小(xiǎo)于 0.01 mg·L-1。
1.3 樣品采集與分(fēn)析測定
1.3.1 樣品采集
養殖試驗結束後将魚饑餓 24 h,以排空其腸道内容物(wù),然後,從每個網箱随機選取 4 尾魚,用 100 mg·L-1間氨基苯甲酸乙酯甲磺酸鹽( MS-222,美國 Sigma 公司) 進行麻醉處理。将魚置于冰袋上,用經肝素鈉抗凝處理的醫用注射器從尾靜脈采血,血樣置于 2 mL 的抗凝管中(zhōng),于 4 ℃、3 000 r·min-1離(lí)心 10 min。将血漿樣品置于-40 ℃ 冰箱中(zhōng)保存,用于血漿生(shēng)化指标的測定。然後,将魚迅速解剖,逐級分(fēn)離(lí)出肝髒、腹脂和腸道并稱質量。将上述樣品用 4 ℃ 預冷後的生(shēng)理鹽水沖洗幹淨後用濾紙(zhǐ)吸幹表面水分(fēn),并快速置于-40 ℃ 冰箱中(zhōng)冷凍保存,用于後續測定工(gōng)作。内髒采集完畢後,用手術刀剝開(kāi)背部皮膚,切取 2 塊肌肉放(fàng)入密封袋中(zhōng),快速置于-40 ℃ 冰箱中(zhōng)冷凍保存,用于肌肉品質的測定。此外(wài),每組保留 2 尾全魚,置于-40 ℃ 冰箱中(zhōng)冷凍保存,用于體(tǐ)組成的測定。
1.3.2 飼料和魚體(tǐ)組成含量測定
将飼料和魚體(tǐ)稱質量後置于培養皿中(zhōng),在 105 ℃ 的烘箱中(zhōng)烘至恒質量後計算得到水分(fēn)含量; 粗蛋白(bái)( N×6.25) 含量采用全自動凱氏定氮儀( FOSS KT260,瑞士 FOSS 公司) 測定;粗脂肪含量采用索氏抽提儀測定; 粗灰分(fēn)采用高溫灼燒法測定; 總能采用氧彈測熱儀( Parr 1281,美國PARR 公司) 測定。
1.3.3 肉品質指标測定
肌肉系水力參照陳代文等的方法進行測定并作适當修改。取 5 g 左右的新鮮背部肌肉, 4 ℃ 懸挂貯存一(yī)段時間後( 保鮮肉) ,稱質量,求出滴水損失。滴水損失的多少反映系水力( water-holding capacity) 大(dà)小(xiǎo)。
滴水損失、蒸煮損失和含肉率等肉品質相關指标的計算公式如下(xià):
滴水損失 = ( 貯存前肉質量-貯存後肉質量) /貯存前肉質量×100%,
蒸煮損失 = ( 蒸煮前肌肉質量-蒸煮後肌肉質量) /蒸煮前肌肉質量×100%,
含肉率 = ( 魚體(tǐ)肉質部分(fēn)質量 /魚體(tǐ)質量) ×100%。
每個網箱随機選取 4 條魚,剝去(qù)背肌部分(fēn)皮膚,用美能達色度計( CR-10, Minolta,日本) 測定肌肉肉色相關指标。
肌肉質構分(fēn)析: 在采樣後 24 h 内,每組選 4 尾魚,各取 1 cm3 背部肌肉,使用質構儀( TA. XT Plus,Stable Micro Systems,英格蘭) 與直徑爲 50 mm 的鋁制壓縮闆進行測定。用 Exponent 軟件( Stable Micro Systems Version 6.0) 記錄測定數據。考察參數主要包括硬度、黏附性、内聚性、咀嚼性以及彈性等,參照Hixson 等的方法進行處理。
1.3.4 血漿生(shēng)理生(shēng)化指标測定
血液生(shēng)化指标中(zhōng)的總蛋白(bái)和白(bái)蛋白(bái)含量均采用南(nán)京建成生(shēng)物(wù)工(gōng)程研究所的試劑盒檢測。其中(zhōng),總蛋白(bái)含量采用考馬斯亮藍(lán)法測定,白(bái)蛋白(bái)含量采用溴甲酚綠比色法測定。補體(tǐ)C3 與 C4 含量和免疫球蛋白(bái) M( IgM) 含量用 ELISA 法測定。血漿髓質過氧化物(wù)酶( MPO) 和溶菌酶活性參照 Zhang 等的方法測定。皮質醇含量參照陳群等的方法測定。
1.3.5 肝髒抗氧化性能測定
準确稱取适量肝髒組織,在冰浴條件下(xià),按照組織與生(shēng)理鹽水質量體(tǐ)積比爲 1 ∶9 的比例混合後進行勻漿,制成 10%( 體(tǐ)積分(fēn)數) 的勻漿液,然後于 4 ℃、3 000 r·min-1 離(lí)心 10 min。取上清液用于超氧化物(wù)歧化酶( SOD) 、過氧化氫酶( CAT) 和谷胱甘肽過氧化物(wù)酶( GSH-Px) 活性以及丙二醛( MDA) 含量和總抗氧化能力( T-AOC) 的測定。樣品上清液中(zhōng)的蛋白(bái)質含量采用考馬斯亮藍(lán)比色法測定。肝髒 SOD、CAT 和 GSH-Px 活性按照 Li 等的方法測定。MDA 含量采用硫代巴比妥酸法測定。T-AOC 參考鞠雪等的方法測定。
1.3.6 腸道消化酶活性測定
準确稱取适量腸道樣品,在冰浴條件下(xià),按照組織與生(shēng)理鹽水質量體(tǐ)積比爲 1 ∶9 的比例混合後進行勻漿,制成 10%的勻漿液,然後于 4 ℃、3 000 r·min-1離(lí)心 10 min。取上清液用于蛋白(bái)酶、脂肪酶、澱粉酶活性的測定。蛋白(bái)酶活性采用福林-酚法測定; 脂肪酶活性參照 Furne 等的方法測定; 澱粉酶活性采用碘-澱粉比色法測定; 各管腸道勻漿樣品中(zhōng)蛋白(bái)含量采用考馬斯亮藍(lán)法測定。
1.4 生(shēng)長性能指标及其計算方法
魚體(tǐ)生(shēng)長性能相關指标: 增重率( WG) 、特定生(shēng)長率( SGR) 、飼料系數( FCR) 、蛋白(bái)效率比( PER) 、氮 /能量保留率( N /ERE) 、肥滿度( CF) 、肝體(tǐ)比( HSI) 、髒體(tǐ)比( VCI) 、腹脂率( IPF) 的計算公式如下(xià):
WG= ( Wt - W0 ) /W0 × 100%; SGR = ( lnWt - lnW0 ) /T × 100%; FCR = F/( Wt - W0 ) ; PER = ( Wt - W0 ) / ( F×CP) ; N/ERE= [( Wt×Ct ) -( W0 ×C0) ] /( F×C) ×100%; CF = Wt /L3 ×100%; HSI = W肝 /Wt ×100%; VCI = W内髒 /Wt×100%; IPF=W腹脂 /Wt×100%。
以上公式中(zhōng): W0 爲魚的初始體(tǐ)質量( g) ; Wt 爲試驗結束時魚的終末體(tǐ)質量( g) ; L 爲試驗結束時魚的體(tǐ)長( cm) ; T 爲養殖時間( d) ; F 爲攝食量( g) ; CP 爲試驗飼料中(zhōng)的蛋白(bái)質含量( %) ; C 爲試驗飼料中(zhōng)的養分(fēn)含量( %) ; C0 爲魚體(tǐ)初始養分(fēn)含量( %) ; Ct 爲養殖結束後魚體(tǐ)的養分(fēn)含量( %) ; W肝、W内髒 和 W腹脂 分(fēn)别表示肝髒、内髒和腹脂質量。
1.5 數據統計與分(fēn)析
采用 SPSS 20.0 軟件對試驗數據進行單因素方差分(fēn)析( one-way ANOVA) ,數據差異顯著時,采用Duncan's檢驗法進行多重比較,差異水平定爲 0.05。試驗結果均保留 2 位小(xiǎo)數,并以 x珋±SE 表示。
2 結果與分(fēn)析
2.1 發酵飼料對鯉魚生(shēng)長性能的影響
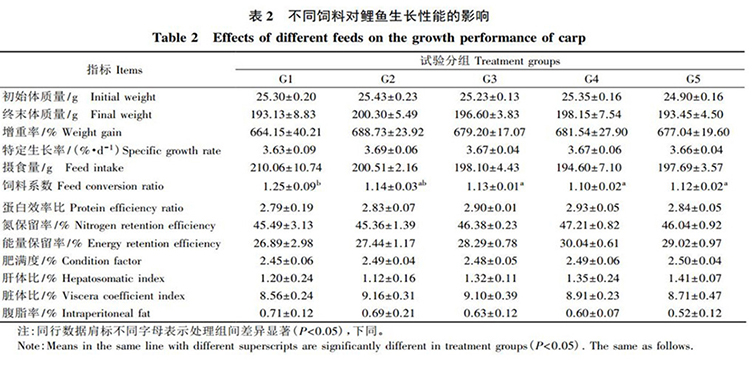
由表 2 可知(zhī): 各組飼料對鯉魚的終末體(tǐ)質量、增重率、特定生(shēng)長率、攝食量、蛋白(bái)效率比、氮保留率、能量保留率、肥滿度、肝體(tǐ)比、髒體(tǐ)比和腹脂率均無顯著影響( P>0.05) ,但顯著影響飼料系數( P<0.05) 。與G1 組相比, G3、G4、G5 組的飼料系數顯著降低, G2 組無顯著變化,而 G3、G4、G5 組間的飼料系數差異不顯著( P>0.05) 。
2.2 發酵飼料對鯉魚體(tǐ)組成的影響
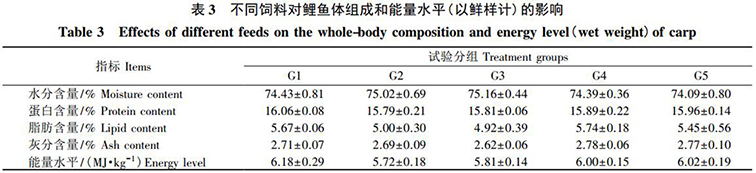
由表 3 可知(zhī): 各組飼料對鯉魚的水分(fēn)、蛋白(bái)質、灰分(fēn)、脂肪含量和能量水平均無顯著影響( P>0.05) 。
2.3 發酵飼料對鯉魚腸道消化酶活性的影響
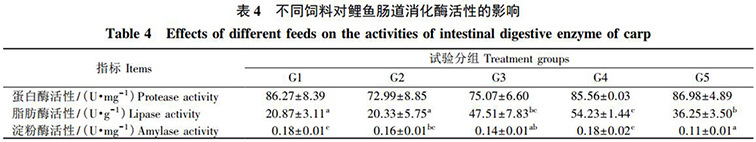
由表 4 可知(zhī): 各組飼料對鯉魚的腸道蛋白(bái)酶活性無顯著影響。G4 組的脂肪酶活性最高,并顯著高于G1、G2 和 G5 組( P<0.05) 。G1 組的澱粉酶活性最高,并顯著高于 G3 和 G5 組( P<0.05) 。
2.4 發酵飼料對鯉魚肌肉品質的影響
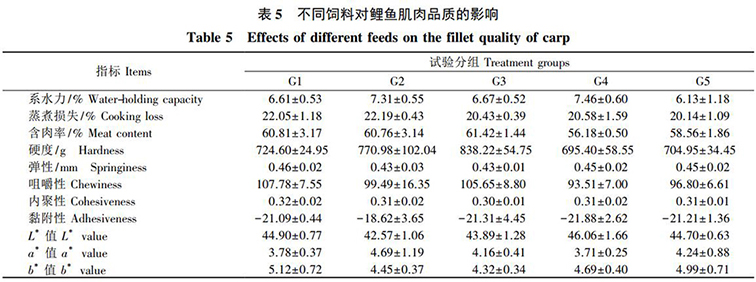
各組飼料對鯉魚的系水力、蒸煮損失、含肉率、硬度、彈性、咀嚼性、内聚性、黏附性以及肉色 a* 、b* 、L* 值均無顯著影響( P>0.05) ( 表 5) 。
2.5 發酵飼料對鯉魚非特異性免疫機能的影響
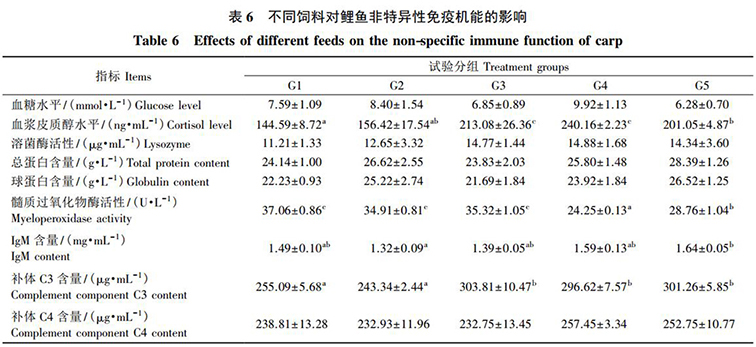
各組飼料對鯉魚的血漿血糖水平、溶菌酶活性以及總蛋白(bái)、球蛋白(bái)和補體(tǐ) C4 含量均無顯著影響( P>0.05) 。各組 IgM 含量均無顯著差異( P>0.05) 。對照組的血漿皮質醇水平和補體(tǐ) C3 含量均與 G2 組之間差異不顯著( P>0.05) ,但卻顯著低于其他各組( P<0.05) 。G1、G2 和 G3 組間的血漿髓質過氧化物(wù)酶活性差異不顯著( P>0.05) ,但均顯著高于 G4、G5 組( P<0.05) ( 表 6) 。
2.6 發酵飼料對鯉魚肝髒抗氧化性能的影響
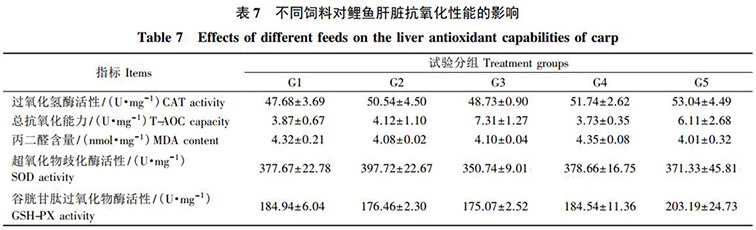
由表 7 可知(zhī): 各組飼料對鯉魚肝髒的 CAT、T-AOC、SOD 和 GSH-PX 活性以及 MDA 含量均無顯著影響( P>0.05) 。
3 讨論
爲了貼近實際生(shēng)産并更真實地反映發酵飼料的使用效果,本試驗在配置飼料時采用了 2 種方法,即直接添加和等蛋白(bái)替代。直接将發酵飼料添加至基礎飼料中(zhōng)的依據在于,在生(shēng)産實踐中(zhōng)絕大(dà)多數企業的飼料配方較爲固定,調整其飼料配方具有較大(dà)的難度,并會在一(yī)定程度上增加飼料生(shēng)産成本。因此,考慮将發酵飼料作爲一(yī)種微量的原料直接添加,具有較強的可操作性。将發酵飼料等蛋白(bái)替代基礎飼料中(zhōng)的菜粕的依據在于,發酵飼料的粗蛋白(bái)水平爲 18%、水分(fēn)爲 42%,烘幹後其蛋白(bái)水平與菜粕最爲接近,用它替代菜粕,不僅能夠滿足與對照組等氮的需要,且對飼料配方的改動較小(xiǎo)。
3.1 發酵飼料對鯉魚生(shēng)長性能和肌肉品質的影響
本試驗中(zhōng),發酵飼料等蛋白(bái)替代 5%菜粕組顯著降低了鯉魚的飼料系數,且這組的攝食量比其他組低。這表明,這組鯉魚的飼料利用率較高,增長單位體(tǐ)質量消耗的飼料量最少。使用這種飼料将顯著降低養殖成本。究其原因,一(yī)方面可能是經過微生(shēng)物(wù)發酵後,飼料中(zhōng)植物(wù)蛋白(bái)源中(zhōng)的抗營養因子含量顯著降低或者被消除; 同時,一(yī)部分(fēn)大(dà)分(fēn)子蛋白(bái)被降解成能夠被水産動物(wù)直接消化吸收的小(xiǎo)分(fēn)子蛋白(bái)、小(xiǎo)肽和氨基酸。經過上述過程,飼料的營養價值得以改善,進而提高了動物(wù)機體(tǐ)的利用率。另一(yī)方面,随着發酵飼料添加或替代量的增加,飼料的酸度會發生(shēng)變化,這在一(yī)定程度上會影響飼料的适口性,進而抑制動物(wù)的食欲,最終造成攝食量下(xià)降。此外(wài),發酵飼料中(zhōng)的益生(shēng)菌可以在動物(wù)機體(tǐ)腸道内生(shēng)長繁衍,促進腸道内維生(shēng)素、蛋白(bái)質和氨基酸等營養成分(fēn)的消化吸收,提高了飼料利用率進而改善了機體(tǐ)的生(shēng)長性能。值得注意的是,發酵飼料等蛋白(bái)替代 5%菜粕組飼料的水分(fēn)含量高于其他組,這也可能對結果造成一(yī)定影響。系水力是最重要的肉品質指标之一(yī),其大(dà)小(xiǎo)經常用滴水損失來描述,滴水損失越多表示系水力越小(xiǎo)。滴水損失越低肉質越好。此外(wài),含肉率的高低也是評價魚類肌肉品質、經濟性狀和生(shēng)産性能的重要指标之一(yī)。而肌肉的顔色( 肉色) 是肉品質優劣的外(wài)觀表現,是人們能夠看到、感覺到的最直觀的印象。質構能反映肉質的軟硬程度和彈性,是肉品食用的主要指标。在本試驗中(zhōng),發酵飼料對肌肉的系水力、蒸煮損失、含肉率、質構以及肉色均無顯著影響。究其原因,一(yī)方面可能是發酵飼料對各組魚體(tǐ)肌肉營養成分(fēn)和蛋白(bái)質結構沒有産生(shēng)顯著影響,從而未對肌肉品質造成影響; 另一(yī)方面,可能是發酵飼料需要添加到一(yī)定量時才會對肌肉品質産生(shēng)顯著影響,而本試驗中(zhōng)發酵飼料的添加量較低,最高的隻有10%,不足以造成顯著影響。這也表明,發酵飼料可以很好地被鯉魚利用,而不會對其肌肉品質産生(shēng)顯著影響。
3.2 發酵飼料對鯉魚消化酶活性的影響
研究表明,腸道是動物(wù)營養物(wù)質消化吸收的主要場所。魚類( 尤其是無胃魚類) 主要靠消化酶進行化學性消化,因此腸道消化酶活性可以在一(yī)定程度上反映魚類的消化能力。本試驗中(zhōng),腸道脂肪酶活性和澱粉酶活性以等蛋白(bái)替代 5% 菜粕組爲最高,等蛋白(bái)替代 10% 菜粕組的脂肪酶和澱粉酶活性均較低,而各組間蛋白(bái)酶活性相差不大(dà)。這表明,發酵飼料可以在一(yī)定程度上提高鯉魚腸道消化酶活性,進而改善其消化機能,但過量的發酵飼料會影響腸道的消化功能。其原因可能是,飼料經發酵後産生(shēng)的有機酸、菌體(tǐ)活性蛋白(bái)、葉酸以及益生(shēng)菌等物(wù)質能有效改善動物(wù)的消化道微生(shēng)态環境,促進消化酶分(fēn)泌,且益生(shēng)菌在增殖過程中(zhōng)會分(fēn)泌一(yī)定的酶類,對内源性酶進行一(yī)定補充,從而提高機體(tǐ)的腸道消化酶活性;也可能是,發酵飼料中(zhōng)抗營養因子的降低或消除,使原料營養成分(fēn)迅速轉化,進而提高動物(wù)機體(tǐ)的消化吸收能力。但是,過量添加發酵飼料會引起腸道内酸度過高,導緻消化酶活性下(xià)降,進而對腸道消化吸收功能造成負面影響。
3.3 發酵飼料對鯉魚免疫機能及肝髒抗氧化能力的影響
本試驗中(zhōng),發酵飼料等蛋白(bái)替代 5% 菜粕組的皮質醇和補體(tǐ) C3 含量均高于對照組,而髓質過氧化物(wù)酶的活力則顯著低于對照組。這可能是發酵飼料和益生(shēng)菌在一(yī)定程度上提高了鯉魚的非特異性免疫機能。這是因爲,血液中(zhōng)血糖和皮質醇水平升高,在一(yī)定程度上反映動物(wù)機體(tǐ)出現了輕微的應激反應; 而補體(tǐ) C3 含量提高,反映發酵飼料能刺激機體(tǐ)的非特異性免疫反應,進而提高其免疫機能; 此外(wài), MPO 活性與中(zhōng)性粒細胞的功能和活力密切相關,其活力下(xià)降反映組織細胞的應激反應減弱。究其原因,一(yī)方面可能是發酵飼料中(zhōng)的小(xiǎo)分(fēn)子蛋白(bái)、肽類和遊離(lí)氨基酸含量相對較高,能夠在一(yī)定程度上增強機體(tǐ)的非特異性免疫力; 另一(yī)方面可能是發酵飼料中(zhōng)的益生(shēng)菌能夠激發機體(tǐ)的體(tǐ)液免疫和細胞免疫來增強機體(tǐ)免疫機能。
細胞在代謝過程中(zhōng)會産生(shēng)大(dà)量的自由基,當其過量時會導緻機體(tǐ)氧化損傷,如細胞膜完整性受損和DNA 結構破壞等。同時,動物(wù)機體(tǐ)内有一(yī)套防禦系統,其由多種抗氧化酶和還原性物(wù)質構成。其中(zhōng), GSH能夠與自由基結合, SOD 可以将 O·-2轉化爲 H2O2,而 GSH-Px 在 GSH 的參與下(xià),催化 H2O2 的分(fēn)解,從而清除自由基。本試驗中(zhōng),發酵飼料對鯉魚肝髒中(zhōng)的 SOD、CAT、GSH-Px、MDA 和 T-AOC 等均無顯著影響。這表明,在飼料中(zhōng)添加發酵飼料和益生(shēng)菌沒有影響鯉魚的抗氧化能力。此試驗結果與王一(yī)娟等和徐奕晴等在中(zhōng)華絨螯蟹上的研究結果不一(yī)緻。究其原因,一(yī)方面可能是益生(shēng)菌和發酵飼料對動物(wù)機體(tǐ)抗氧化能力的調節作用存在種間特異性,且與蛋白(bái)源的替代水平及動物(wù)養殖周期的長短等因素密切相關; 另一(yī)方面可能是本試驗中(zhōng)發酵飼料添加量相對較低,不足以影響機體(tǐ)的抗氧化功能。
本試驗結果表明,發酵飼料等蛋白(bái)替代 5%菜粕組的飼料系數較低、腸道消化酶活性較高且非特異性免疫機能較強,而其蛋白(bái)效率比及氮保留率均與對照組相差不大(dà),表明這組飼料不僅可以提高魚體(tǐ)對飼料的利用率,還可以在一(yī)定程度上改善魚體(tǐ)的消化機能和免疫力。在實際生(shēng)産中(zhōng),使用這組飼料來飼喂鯉魚,将會在一(yī)定程度上降低飼料消耗量并改善魚體(tǐ)抗病力,進而降低養殖成本并改善養殖效益,也不會影響鯉魚的肌肉品質。
參考文獻(略)。
新聞中(zhōng)心
關于我(wǒ)(wǒ)們
産品中(zhōng)心
文章發表
技術支持
聯系我(wǒ)(wǒ)們

地址:南(nán)京市江甯開(kāi)發區東吉大(dà)道1号
電話(huà) : 025-52776616
網址:www.njxjz.com.cn